Anticuerpo monoclonal de conejo Bcl-6 (3J5)
Conjugación: No conjugado
Anticuerpo monoclonal de conejo recombinante
Aplicación
Reactividad
Humano
Nombre del gen
BCL6
Almacenamiento
Alicuotar y almacenar a -20 °C (válido por 12 meses). Evitar ciclos de congelación y descongelación.
Resumen
| Nombre del producto | Anticuerpo monoclonal de conejo Bcl-6 (3J5) |
| Descripción | Anticuerpo monoclonal de conejo recombinante |
| Hospedador | Conejo |
| Reactividad | Humano |
| Conjugación | No conjugado |
| Modificación | Sin modificar |
| isotipo | IgG |
| Clonalidad | Monoclonal |
| Forma | Líquido |
| Concentración | No conjugado |
| Almacenamiento | Alicuotar y almacenar a -20 °C (válido por 12 meses). Evitar ciclos de congelación y descongelación. |
| Envío | Bolsas de hielo. |
| Buffer | IgG de conejo en solución salina tamponada con fosfato, pH 7,4, 150 mM de NaCl, 0,02 % de conservante de nuevo tipo N y 50 % de glicerol. Conservar a +4 °C a corto plazo. Conservar a -20 °C a largo plazo. Evitar el ciclo de congelación/descongelación. |
| Purificación | Purificación por afinidad |
Información del antígeno
| Nombre del gen | BCL6 |
| Nombres alternativos | B-cell lymphoma 6 protein; BCL-6; B-cell lymphoma 5 protein; BCL-5; Protein LAZ-3; Zinc finger and BTB domain-containing protein 27; Zinc finger protein 51; BCL6; BCL5; LAZ3; ZBTB27; ZNF51 |
| Gene ID | 604 |
| SwissProt ID | P41182 |
| Immunogen | Proteína recombinante de BCL-6 humana |
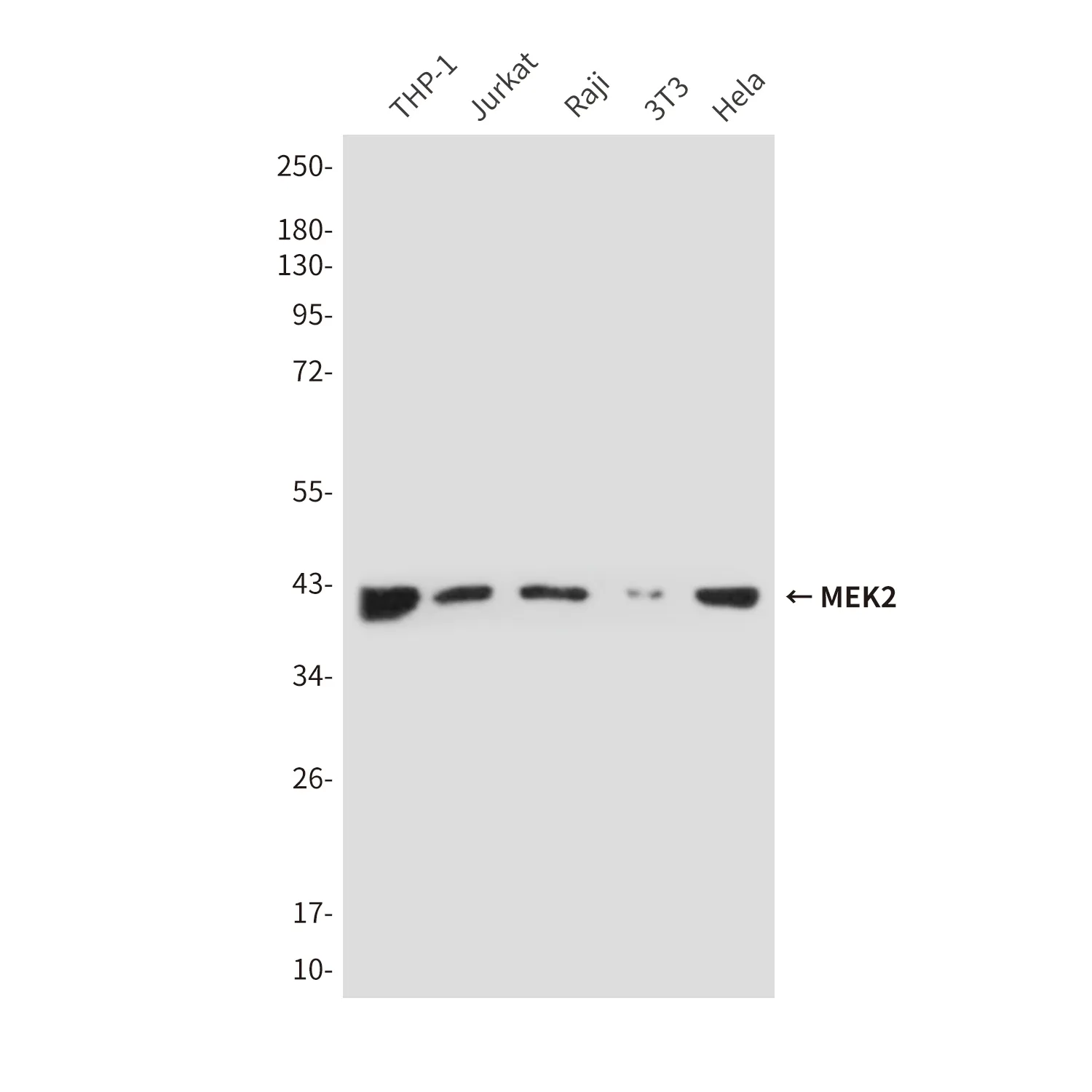
Aplicación
| Aplicación | WB,IHC,IF-P |
| Proporción de dilución | WB 1:1000-1:5000,IHC 1:200-1:2000,IF-P 1:200-1:2000 |
| Peso molecular | 79kDa |
Área de investigación
| Epigenetics and Nuclear Signaling |
Antecedentes
| Bcl-6, un represor transcripcional, se une a elementos de ADN similares al reconocimiento de Stat e influye en el desarrollo del centro germinal y la diferenciación celular. Además, Bcl-6 regula negativamente la expresión de NFκB, inhibiendo así las funciones celulares mediadas por NFκB. La acetilación de Bcl-6, dependiente de HDAC y del regulador de información silenciosa (SIR)-2, provoca una regulación negativa de la actividad al inhibir la capacidad de Bcl-6 para reclutar complejos que contienen histonas desacetilasas (HDAC). Este represor transcripcional, necesario principalmente para la formación del centro germinal (GC) y la maduración de la afinidad de anticuerpos, presenta diferentes mecanismos de acción específicos del linaje y las funciones biológicas. Forma complejos con diferentes correpresores e histonas desacetilasas para reprimir la expresión transcripcional de diferentes subconjuntos de genes diana. Reprime sus genes diana uniéndose directamente a la secuencia de ADN 5'-TTCCTAGAA-3' (sitio de unión a BCL6) o indirectamente reprimiendo la actividad transcripcional de los factores de transcripción. En las células B del GC, reprime genes que funcionan en la diferenciación, inflamación, apoptosis y control del ciclo celular, también autorregula su expresión transcripcional y aumenta, indirectamente, la expresión de algunos genes importantes para las reacciones del GC, como AICDA, a través de la represión de la expresión de microARN, como miR155. Una función importante es permitir que las células B del GC proliferen muy rápidamente en respuesta a antígenos dependientes de células T y toleren las roturas fisiológicas del ADN requeridas para la recombinación de cambio de clase de inmunoglobulina y la hipermutación somática sin inducir una respuesta apoptótica dependiente de p53/TP53. En las células T CD4(+) auxiliares foliculares (células T(FH)), promueve la expresión de genes relacionados con T(FH) pero inhibe la diferenciación de células T(H)1, T(H)2 y T(H)17. También es necesario para el establecimiento y mantenimiento de la memoria inmunológica tanto para células T como B. Suprime la proliferación de macrófagos a través de la competencia con STAT5 por motivos de unión a STAT que se unen a ciertos genes diana, como CCL2 y CCND2. En respuesta al estrés genotóxico, controla la detención del ciclo celular en células B GC de maneras dependientes e independientes de p53/TP53. Además, también controla la neurogénesis a través de la alteración de la composición de complejos transcripcionales dependientes de NOTCH en dianas NOTCH selectivas, como HES5, incluido el reclutamiento de la desacetilasa SIRT1 y resultando en un silenciamiento epigenético que conduce a la diferenciación neuronal. |